

Ochenta y cuatro años atrás, Weindling quien, en 1932, describía por primera vez la capacidad de Trichoderma para parasitar las hifas del fitopatógeno Rhizoctonia solani, sentando las bases del concepto de micoparasitismo.
Tuvieron que pasaron trece años para que en 1945 Brian y McGowan documentaran el antagonismo de Trichoderma frente a Phoma betae mediante cultivos duales en placa, demostrando también la presencia de antibiosis.
Desde entonces, la investigación en torno a este género no ha dejado de crecer, hasta convertirlo hoy en el agente de control biológico de hongos y oomicetos más comercializado en el mundo.
En España, el grupo del Dr. Enrique Monte en la Universidad de Salamanca, lleva más de cuatro décadas —desde la tesis doctoral de Monte en 1986— contribuyendo de manera decisiva al conocimiento de Trichoderma. Su trabajo abarca la caracterización morfológica, molecular y genómica de cepas, la elucidación de los mecanismos de biocontrol, las interacciones con las plantas y los nuevos roles como bioestimulante.
La primera patente biotecnológica de la Universidad de Salamanca fue precisamente una formulación de Trichoderma desarrollada por este grupo, que también registró el primer biofungicida comercial en España —TUSAL® (Trichoderma de la Universidad de Salamanca)— y uno de los primeros cuatro resgitrados en la Unión Europea.
En el contexto actual, la presión regulatoria sobre los plaguicidas de síntesis química, la creciente resistencia de los patógenos, los objetivos de sostenibilidad de la agricultura y la demanda de alimentos inocuos han impulsado de forma extraordinaria el mercado global de los productos biológicos.
En América Latina, donde la diversidad de cultivos, la presión de patógenos y las condiciones climáticas representan un desafío permanente, Trichoderma he sobresalido como una solución polivalente.
Y es que Trichoderma no solo controla enfermedades, sino que estimula el crecimiento vegetal, mejora la estructura del suelo y modula el microbioma de la rizosfera. Científicos como Woo, Hermosa, Lorito y Monte sintetizaron esta perspectiva en una revisión publicada en Nature Reviews Microbiology (2023), donde se consolidaba a Trichoderma como el microorganismo beneficioso multifuncional por excelencia para la agricultura ecosostenible.
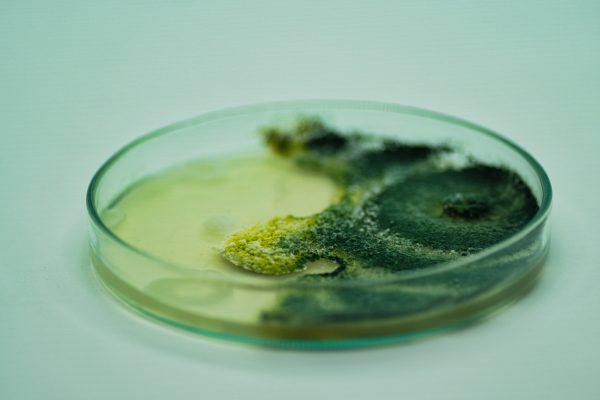
Crecimiento de Trichoderma spp. en una placa de Petri con agar de dextrosa de patata (PDA), antagonismo y antibióticos con Bacillus subtilis, efecto antibiótico y Paecilomyces lilacinus.
DIVERSIDAD TAXONÓMICA Y CARACTERIZACIÓN MOLECULAR
El género Trichoderma (orden Hypocreales, familia Hypocreaceae) comprende actualmente más de 460 especies descritas, número que ha crecido exponencialmente en las últimas dos décadas gracias a la incorporación de herramientas moleculares. Esta expansión taxonómica ha exigido definir criterios moleculares robustos para la identificación de especies, superando las limitaciones del diagnóstico morfológico tradicional.
Genes marcadores de referencia: El grupo de Monte y Hermosa fue pionero en aplicar la secuenciación génica a la caracterización de cepas de Trichoderma con potencial de biocontrol. Trabajos realizados entre 1997 y 2004 establecieron el uso combinado de la región ITS1 del ADNr y el gen tef1 (factor de elongación de la traducción 1-alfa) como marcadores taxonómicos de referencia, logrando distinguir seis nuevos genotipos en T. harzianum e identificar por primera vez T. aggressivum genotipo Th2 en Europa.
En la actualidad, la comunidad científica internacional ha consensuado tres marcadores principales para la delimitación de especies en Trichoderma: la región ITS, el gen tef1 y el gen rpb2 (que codifica la subunidad 2 de la ARN polimerasa II). El gen acl1 (ATP citrato liasa 1) se emplea como complemento.
En este sentido, el grupo de Salamanca alerta contra lo que denominan ‘vandalismo taxonómico’: la proliferación de descripciones de nuevas especies sin el uso riguroso de estos tres marcadores en combinación, lo que genera confusión en la nomenclatura y dificulta la reproducibilidad de los ensayos de biocontrol.
El análisis comparativo de 33 genomas de Trichoderma ha permitido organizar el género en clados bien definidos: Harzianum, Virens y Brevicompactum, con las secciones Longibrachiatum y Trichoderma, además de varios linajes más basales. Este árbol filogenómico es esencial para entender la distribución de las capacidades de biocontrol.
Importancia de la cepa en el biocontrol: El biocontrol con Trichoderma es un ‘desarrollo a medida’. Cada especie, e incluso cada cepa dentro de una especie, produce una colección diferente de enzimas hidrolíticas y metabolitos secundarios, lo que determina su espectro de actividad frente a los patógenos diana. La selección de la cepa adecuada para cada cultivo, patosistema y condición agroclimática es, por tanto, un paso crítico previo a cualquier aplicación comercial.

Las quitinasas de T. viride han mostrado actividad frente a la quitina de la matriz peritrófica del gusano de seda Bombyx mori.
TRICHODERMA Y SUS TRES HABILIDADES COMERCIALES
Desde una perspectiva aplicada, Trichoderma es comercializado por tres habilidades fundamentales, que frecuentemente actúan de forma simultánea y sinérgica:
- Agente de control biológico directo: micoparasitismo, antibiosis y competición con los patógenos.
- Bioestimulante: promoción del crecimiento vegetal y desarrollo del sistema radical, mediada por la producción de ácido indolacético (AIA), actividad ACC desaminasa y señalización hormonal.
- Biocontrol indirecto (‘priming’): inducción sistémica de resistencia (ISR) frente a enfermedades, plagas y estreses abióticos, a través de la activación de las rutas de señalización del ácido salicílico (SA), ácido jasmónico (JA) y etileno (ET).
Esta triple funcionalidad sitúa a Trichoderma en una posición única dentro del mercado de productos biológicos, diferenciándolo de otros agentes de biocontrol con espectros de acción más restringidos como:
Trichoderma está organizado en cinco grandes categorías: micoparasitismo; antibiosis; control enzimático y por metabolitos secundarios frente a insectos y nematodos; competición en suelo, rizosfera y endosfera; y atracción de enemigos naturales de las plagas mediante compuestos orgánicos volátiles (VOCs). A continuación se analiza cada una de estas categorías.
Micoparasitismo: es el mecanismo más estudiado y probablemente el más relevante en el biocontrol de hongos y oomicetos fitopatógenos. Se trata de una simbiosis antagónica en la que Trichoderma reconoce, ataca, penetra y destruye las hifas de otros hongos mediante una secuencia de eventos altamente coordinada.
El proceso comprende varias etapas: (i) reconocimiento quimiotrópico del hospedador, que implica la detección de señales químicas emitidas por el patógeno; (ii) contacto y coiling de las hifas de Trichoderma en torno a las hifas del hospedador; (iii) secreción de enzimas hidrolíticas extracelulares que degradan la pared celular fúngica; y (iv) penetración y lisis del contenido citoplasmático del patógeno.
A través del micoparasitismo, Trichoderma ha incorporado a su propio genoma genes robados a sus presas fúngicas, un fenómeno de transferencia horizontal de genes que ha ampliado el arsenal enzimático del género a lo largo de la evolución.
Trichoderma es eficaz contra prácticamente todas las especies del género Phytophthora, incluyendo P. cinnamomi, P. infestans y otras, así como contra Fusarium, Rhizoctonia, Botrytis, Sclerotinia, Pythium, Verticillium y otros fitopatógenos de importancia agrícola global. En ensayos documentados por el grupo de Salamanca, T. asperellum y T. ghanense demostraron efectividad como suplemento del compost en el control de Fusarium oxysporum f. sp. lycopersici (FOL) y F. oxysporum f. sp. radicis-lycopersici (FORL) en tomate cultivado en Argelia.
El Arsenal Enzimático de Trichoderma: Trichoderma posee el mayor arsenal de enzimas degradadoras de la pared celular (CWDEs, Cell Wall-Degrading Enzymes) de todo el Reino Fungi, comprendiendo tres grandes familias enzimáticas:
Glucanasas (beta-1,3-glucanasas y beta-1,6-glucanasas): degradan los glucanos que forman el esqueleto estructural de la pared celular de la mayoría de los hongos fitopatógenos.
Quitinasas: hidrolizan la quitina, componente mayoritario de la pared celular de los hongos y del exoesqueleto de los insectos.
Proteasas: degradan las proteínas estructurales de la pared celular fúngica y, en algunos casos, interfieren directamente con la actividad enzimática de los patógenos.
El grupo del Dr. Monte caracterizó molecularmente 7 enzimas clave y sus correspondientes genes en T. harzianum, demostrando que el sobrenadante de cultivo de T. harzianum —es decir, sin micelio— es capaz por sí solo de degradar la pared celular de Botrytis cinerea de forma progresiva y completa en siete horas.
Un hallazgo fundamental de estos estudios es que cada cepa de Trichoderma produce una colección distinta de hidrolasas, y que la combinación de enzimas es determinante para la eficacia del biocontrol.
Control de nematodos: los fitonematodos, en particular los del género Meloidogyne (nematodos formadores de nódulos radiculares), representan una de las principales plagas agrícolas a nivel global, con pérdidas anuales estimadas en decenas de miles de millones de dólares. El grupo de Monte y Hermosa ha documentado la actividad nematicida de Trichoderma mediante dos mecanismos complementarios:
Acción enzimática directa: las quitinasas y proteasas de Trichoderma, especialmente la proteasa PRA1, son capaces de parasitar y destruir los huevos de Meloidogyne. La quitina presente en la cubierta del huevo y en la cutícula del nematodo es el sustrato de las quitinasas, mientras que las proteasas atacan las proteínas estructurales (Suárez et al., 2004).
‘Priming’ de defensas vegetales frente a nematodos: Estudios científicos demostraron en un elegante sistema de raíz dividida (‘split-root’) que T. atroviride es capaz de inducir resistencia sistémica frente a Meloidogyne javanica.

T. atroviride produce los metabolitos volátiles 6-PP y 1-octen-3-ol, cuya aplicación en la dieta de larvas de Spodoptera frugiperda genera función repelente.
SU PODER PARA EL CONTROL DE PLAGAS
El control de insectos representa una de las fronteras más innovadoras de la investigación en Trichoderma. Los mecanismos descritos incluyen:
Acción enzimática sobre insectos: Las quitinasas de T. viride han mostrado actividad frente a la quitina de la matriz peritrófica del gusano de seda Bombyx mori (Berini et al., 2016), abriendo la vía al uso de estas enzimas en el control de lepidópteros.
Asimismo, se ha documentado acción insecticida sobre Aedes aegypti, mosquito vector de enfermedades tropicales como el dengue, el zika y la fiebre amarilla, lo que sugiere potenciales aplicaciones en salud pública más allá de la fitoprotección.
VOCs con función repelente: trabajos de investigación demostraron que T. atroviride produce los metabolitos volátiles 6-PP y 1-octen-3-ol, cuya aplicación en la dieta de larvas de Spodoptera frugiperda —el gusano cogollero de maíz y algodón, una de las plagas más devastadoras de América Latina— genera función repelente, menor desarrollo larvario y más de un 50% de mortalidad. Esta acción se ejerce directamente sobre el insecto, a través de la dieta, representando un mecanismo de bioinsecticida basado en metabolitos volátiles.
Modificación del microbioma intestinal de los insectos: es un mecanismo emergente. En 2023, Di Lelio se publicó un publicaron en PNAS un hallazgo de enorme relevancia conceptual: el tratamiento de semillas de tomate con Trichoderma modifica el metaboloma de la planta, y este cambio en la composición química de las hojas altera a su vez el microbioma bacteriano intestinal de Spodoptera littoralis cuando se alimenta de dichas hojas. Específicamente, se observó una menor abundancia de la bacteria dominante Enterococcus casseliflavus en el intestino de las larvas, lo que se traduce en menor supervivencia, desarrollo y longevidad del insecto.
VOCs como atrayentes de parasitoides y predadores: Más allá de su acción directa sobre los insectos, los VOCs de Trichoderma desempeñan un papel ecológico de gran interés: la atracción de parasitoides y predadores naturales de las plagas.
En 2018 se demostró que que T. atroviride, al colonizar la raíz del maíz, produce VOCs que se difunden a través del suelo y atraen a Campoletis sonorensis, un parasitoide himenóptero de las larvas de Spodoptera frugiperda. El número de insectos parasitados aumentó significativamente cuando T. atroviride producía la cantidad adecuada de 6-PP, evidenciando una cadena trófica de control biológico mediada por este hongo.

Tomate afectado por el ataque de Phytophthora infestans.
COMPETICIÓN EN SUELO, RIZOSFERA Y ENDOSFERA
Para ejercer su función de biocontrol, Trichoderma debe establecerse y persistir en el suelo, compitiendo con la microbiota autóctona y resistiendo las condiciones ambientales adversas. El grupo de los doctores Monte y Hermosa ha estudiado los mecanismos que confieren a Trichoderma ventajas competitivas en este entorno:
Resistencia al calor: la cepa T. harzianum T34, ampliamente utilizada como ACB, mostró capacidad de adquirir termotolerancia mediante la expresión heteróloga de la ‘heat shock protein’ HSP23 de T. virens T59 (Montero-Barrientos, 2007). Asimismo, la sobreexpresión del gen hsp70 en T. harzianum T34 confirió resistencia simultánea al calor (choque térmico a 45°C), el estrés osmótico y el estrés oxidativo, sin comprometer la capacidad de biocontrol de la cepa (Montero-Barrientos, 2008). Estos hallazgos tienen implicaciones prácticas directas para el desarrollo de cepas de Trichoderma adaptadas al uso en solarización de suelos y en condiciones de temperatura elevada.
Resistencia al estrés oxidativo: En la rizosfera, Trichoderma debe hacer frente a los radicales libres de oxígeno (ROS). La sobreexpresión de la tiorredoxina TvDim1 de T. virens aumenta significativamente la biomasa fúngica tras un estrés oxidativo con H₂O₂ 3 mM, mientras que los mutantes silenciados para este gen no crecieron en dichas condiciones. Por su parte, la NADPH oxidasa (NOX1) de T. harzianum regula la producción de ROS propios, incrementa las actividades proteasa, quitinasa y celulasa, y mejora el biocontrol de Pythium ultimum.
Competición en la rizosfera y modificación del microbioma: Trichoderma no solo compite con los patógenos por espacio y nutrientes, sino que actúa como modulador activo del microbioma rizosférico. Al acidificar el suelo en su entorno, Trichoderma provoca cambios en la abundancia relativa y en la composición taxonómica de las comunidades de bacterias y hongos del suelo.
La capacidad de orquestar el microbioma rizosférico hacia una comunidad más favorable para la planta representa un mecanismo de biocontrol de orden superior, con efectos potencialmente persistentes en el tiempo y en el espacio. Desde la perspectiva agronómica, implica que la aplicación de Trichoderma tiene un efecto ecosistémico que trasciende la acción directa sobre el patógeno diana.
Competición en la endosfera: Uno de los hallazgos más relevantes del grupo de Monte y Hermosa en los últimos años es la capacidad de T. harzianum para colonizar la raíz de olivo de forma endofítica, estableciéndose en el interior del tejido radical sin causar daño a la planta. Esta colonización endofítica tiene consecuencias fundamentales en el control de Verticillium dahliae (Vd), el principal patógeno vascular del olivo en la cuenca mediterránea.
La investigadora Irene Carrero-Carrón demostró que T. harzianum, una vez establecido como endófito, dificulta físicamente el acceso de V. dahliae a los haces vasculares de los plantones de olivo, previniendo el movimiento sistémico del patógeno dentro de la planta. La estrategia propuesta por el grupo de Salamanca es de notable ingenio agronómico: producir plantones de olivo sobre pies de acebuche (el olivo silvestre, Olea europaea var. sylvestris) previamente colonizados por Trichoderma, aprovechando la resistencia natural del acebuche como portainjerto y la protección endofítica de Trichoderma de forma combinada.
Además de la protección frente a patógenos vasculares, la colonización endofítica promueve el crecimiento de la planta, mejora la tolerancia a estreses abióticos como la sequía y la salinidad, e induce defensas sistémicas frente a patógenos aéreos.

El grupo de Monte y Hermosa ha documentado la actividad nematicida de Trichoderma.
BIOCONTROL INDIRECTO: ‘PRIMING’ E INDUCCIÓN SISTÉMICA DE RESISTENCIA (ISR)
Concepto y rutas de señalización: El ‘priming’ de defensas vegetales por Trichoderma es un mecanismo de biocontrol indirecto de enorme relevancia agronómica. Este actúa sobre la planta, preparando su sistema inmune para responder de forma más rápida y eficaz ante la llegada de un patógeno, plaga o estrés abiótico. La defensa no se activa de forma constitutiva (lo que tendría costes metabólicos elevados), sino que queda ‘en estado de alerta’ y se dispara plenamente solo cuando llega el reto biótico o abiótico.
Los mecanismos moleculares del ‘priming’ mediado por Trichoderma implican la activación diferencial de las rutas del ácido salicílico (SA), el ácido jasmónico (JA) y el etileno (ET), según la cepa de Trichoderma aplicada, el inóculo utilizado y el genotipo de la planta huésped. La ruta del SA es primordialmente efectora frente a patógenos biotrófos, mientras que la del JA/ET es más activa frente a necrotrófos y herbívoros. El hecho de que Trichoderma pueda activar ambas rutas —o modularlas de forma específica— le confiere una versatilidad defensiva notable, según un trabjao de la Dra. Rosa Hermosa y María Morán-Diez.
Las moléculas efectoras que desencadenan el ‘priming’ incluyen proteínas secretadas por Trichoderma al apoplasto radical —denominadas efectores— que son reconocidas por la planta como señales de activación de la ISR.
Demostración experimental del priming: En Costa Rica, el trabajo de William Rivera-Méndez documentó la efectividad del priming de T. asperellum en condiciones de campo, en el control de Sclerotium cepivorum (agente de la podredumbre blanca) en cebolla tropical en Costa Rica. Los análisis moleculares revelaron que T. asperellum induce la producción de SA a 1 y 7 días tras la aplicación, y que el estado de ‘priming’ desaparece a los 21 días a nivel de metabolitos, pero queda guardado en la ‘memoria transcripcional’ de la planta. Este concepto de memoria epigenética del ‘priming’ es de gran relevancia para el diseño de protocolos de aplicación: la frecuencia y el momento del tratamiento con Trichoderma deben optimizarse para garantizar que el estado de priming esté activo en el momento de máximo riesgo de infección.
La demostración más elegante de la naturaleza sistémica del ‘priming’ fue la que se publicó en el trabjo de Medeiros en 2017 en el sistema ‘split-root’ frente a M. javanica, ya descrita en el apartado de control de nematodos. Este experimento evidenció que Trichoderma no necesita estar en contacto físico con el patógeno para ejercer su función de biocontrol: su presencia en una zona del sistema radical es suficiente para conferir protección sistémica en otras zonas de la planta.
Interacción multitrófica: El trabajo publicado por Di Lelio en PNAS representa probablemente el mecanismo más novedoso y conceptualmente más disruptivo descrito hasta la fecha para el biocontrol mediado por Trichoderma. Su aplicación a semillas de tomate indujo cambios en el metaboloma de las hojas de la planta —es decir, en la composición de sus metabolitos secundarios— sin alterar su apariencia macroscópica. Cuando las larvas de S. littoralis se alimentaron de estas hojas, su microbioma intestinal fue perturbado, con una notable reducción en la abundancia de Enterococcus casseliflavus. Esta alteración del microbioma intestinal se tradujo en menor tasa de supervivencia, menor desarrollo y reducida longevidad del insecto.
Este mecanismo implica una cadena de eventos interspecífica de cuatro actores: Trichoderma → planta → hoja → insecto → microbioma del insecto, lo que lo convierte en el primer ejemplo documentado de biocontrol de insectos mediado por la modificación de la interacción planta-microbioma intestinal del herbívoro. El Dr. Monte señaló en su comentario en PNAS que este hallazgo abre una nueva categoría de biocontrol y refuerza el argumento de que Trichoderma actúa como orquestador de ecosistemas, mucho más allá de ser un simple parásito de hongos.
